| GISdevelopment.net ---> AARS ---> ACRS 2000 ---> Landuse |
Development of Trunk-Canopy
Biomass and Morphology Indices from Quadpolarized Radar Data
Peter N. Tiangco and Bruce
C. Forester
School of Geomatic Engineering
University of New South Wales Syney, NSW 2052, Australia
Email: z2146531@student.unsw.edu.au, B.forster@unsw.edu.au
AbstractSchool of Geomatic Engineering
University of New South Wales Syney, NSW 2052, Australia
Email: z2146531@student.unsw.edu.au, B.forster@unsw.edu.au
The capability of microwave energy to penetrate forest vegetation makes possible the extraction of information on both the crown and trunk components from radar data. At C-band, the backscattered energy is correlated mainly with the crown constituents such as the leaves, twigs and small branches. Information on the other components beneath the canopy can be sensed through the use of bands with longer wavelengths such as the L-or P-band. The sensitivity of co-polarized and cross-polarized waves to the shapes and orientation of different tree constituents provide an added advantage in the information extraction procedure. The Trunk-Canopy Biomass Index (TCBI), which is the sum of the L-HH and C-HV backscatter, can be a measure of the total aboveground biomass as both the crown and trunk layers are taken into consideration. Owing to possible morphological variation, the relationship between TCBI and biomass is however not expected to be unique for a whole forest vegetation. It is important therefore that stand structure be first considered to allow a more accurate biomass assessment by the TCBI. An index of the relative proportions of the crown and trunk may be indicative of the approximate tree morphology. It is believed that the Trunk-Canopy Morpholoyg Index (TCMI), which is the ratio of the L-HH to C-HV backscatter, provides a measure of tree structure. In this study, two categories are used to classify stands according to structure: the needle-leaved pines/conifers and the broad-leaved deciduous/evergreen trees. A two stage procedure of forest biomass estimation is therefore proposed. The first stage involves the determination of the stand structure category based on the TCMI. Once the structure is known, a specific structure-dependent TCBI could then be applied for the biomass estimation process. The effectiveness of these indices is assessed by applying them to actual and modeled data interpolated from published works of other investigators. Stand structure and total aboveground biomass were found to be highly correlated with TCMI and TCMI and TCBI, respectively. Comparison of the results is made difficult by the limitations in the amount of data available from the published studies and the possible errors introduced during the interpolation process. In order to verify the validity of these results, further application of these indices using AIRSAR images and adequate amount of actual and measured values from an independent study site in the Blue Mountains area in New South Wales, Australia will be conducted.
1. Introduction
The importance of quantifying and monitoring forest vegetation, given the vital productive, protective, and regenerative functions of this natural resource, is undeniable. Information on the amount and extent of vegetation provides an insight on what or how much to expect from a specific forest area in terms of these functions. From another perspective, the information can be useful in determining whether or not forest rehabiliation or other appropriate actions are needed in consideration of these three major functions.
Forest aboveground biomass, or the quantity of vegetative material per unit area, is one of the parameters recognized as a good indicator of forest condition. The use of radar remote sensing to
Provide estimates of this parameter has been receiving increasing over recent years. This is mainly due to ability of the radar to provide data independent of solar illumination and weather conditions. In addition, radar waves, depending n the wavelength/frequency, can be employed to scope different sections of the vegetation profile. Information on crown layer components (e.g. foliage) can be inferred from the radar backscatter at high frequency bands such as C-and X-while information pertaining to the trunks and lower branches can be obtained through the longer and more penetrative wavelengths of L-and P-bands.
Since the backscattering coefficient is influenced not only by the quantity of biomass but by how the individual components are oriented and distributed throughout the entire tree length, it is also possible to determine the general tree/stand structure based on the radar data. In fact, due to possible morphological variations between stands within a given forest area, it is recommended that the determination of the general tree/stand structure must precede biomass estimation and that radar data-based biomass equations should be formulated based on the structure. This is to avoid erroneous results will most likely be produced as it is possible to obtain different backscatter readings from two stands containing the same amount of biomass but are structurally-different.
2. Synergism of Radar Parameters for Optimum Information Extraction
The amount and quality of information which can be inferred from radar data depend on the characteristics of the target and the radar system. Important factors under the former relate to the roughness, geometric and dielectric properties of the imaged surface while under the latter are the microwave frequency/wavelength, polarization and incidence angle used in the data acquisition process. Accurate inference of vegetation properties results from the formulation of a well-established relationship between the target and the radar parameters. Being well-established implies present and a clear understanding of how these mechanisms relate to the radar parameters.
The advent of multi-parameter imaging radars extricated investigators from the limitations of using mono-band radar data in carrying out forest-related research. The opportunity lies in the simultaneous use of the radar parameters for optimum extraction of information from radar imagery. It is believed that a better assessment of forest resources could be achieved by utilizing a combination of the parameters. The theoretical basis for this premise and the different combinations will be presented in the following topics. Focus will be on the more controllable radar system-based components such as wavelength and polarization with forest aboveground biomass and stand structure assessments as the particular areas of application.
2.1 Forest aboveground biomass assessment
2.1.1 Combined wavelength and polarization estimation
The total forest aboveground biomass is the summation of the biomass of the crown and bole components of all the trees in the area under consideration. Obviously, to derive an estimate of this quantity from radar data, information on both the crown and the bole should be available. A simple model of the total biomass could then be given as
Btotal = Bt + Bc (1)
Where Btotal represents the total aboveground biomass, Bt the trunk biomass, and Bc the crown biomass.
The C-band, with its relatively shorter wavelength, is sensitive to the upper layer of the vegetation such as the leaves, twigs and small branches of the crown. The more penetrative wavelength of L-band can pass through a greater volume of the canopy and interacts with bigger structures at the lowermost canopy portion and more regularly, with the trunk and big branches. Most of the returns from the latter is from tree-ground double bounce backscattering which occurs mainly due to the Vertical orientation of the trunk with respect to the ground hence forming a corner a corner reflector-like structure. Thus,
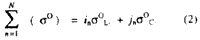
where
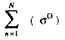 is the backscattering coefficient sum
correlated with the total aboveground biomass;
is the backscattering coefficient sum
correlated with the total aboveground biomass; i and j are the unit vectors in x and y axes in the xy coordinate system corresponding to the backscattering coefficients of the L-and C-band, respectively.
The type of polarization employed determines the radar response to the various shapes and orientations of the scattering mechanisms within the canopy. Backscatter from cross-polarized waves tends to be related to the canopy volume rather than the lower components such as the soil and as such may be an indicator of crown biomass (ESA, 1995). Incoming vertically-polarized waves readily interact with the vertical components of the canopy. Due to the nature of its orientation, horizontally-polarized radiation tends to have a deeper degree of penetration and is less likely to be affected by canopy attenuation. The highest correlations between radar backscatter and total biomass are obtained using waves with HV-and HH- polarization: VV- waves are more sensitive to the components of the crown and tend to saturate at lower total biomass levels (Dobson et al., 1995a).
Integrating the theories pertaining to the sensitivities of the different radar wavelengths and polarizations, we can deduce the following relationships:
so (Bt+ib) " so L-HH (3)
so (BL+b) " so C-HV (4)
where Bt+ib refers to the biomass of the trunk and lower branches, BL+b is the biomass of the leaves and most of the brances, while soL-HH and s C-HV are the L-HH and C-HV backscattering coefficients, respectively. Considering the above relationships, equation (2) could then be written as
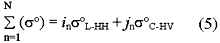
Where isoL-HH + jsoC-HV is called the Trunk-Canopy Biomass Index (TCBI). To avoid possible overestimation of total aboveground biomass, other polarization combinations of the L-and C-bands wee not included in the equation as the biomass values they represent are already included in the soL-HH and soC-HV backscatter.
2.1.2. Saturation of radar measurements
The amount of density of the scattering components within the crown, in addition to wavelength and dielectric constant, also influences the penetration depth achieved by the transmitted energy. The influence stems from the changes in path length and the degree of attenuation imposed by the crown components on the incident radiation. As the energy travels through the canopy, it is gradually depleted by the scattering caused by dielectric discontinuities in the different canopy layers such that there is less energy available to the next layer (UNSW course notes, 1996). If we assume a continuous increase in the dimensions and amount of canopy components from left to right of a forest profile, there will be a point in the profile wherein even the longer wavelength of the L-band would no longer be able to pass through the crown layer due to attenuation.
The increase in the dimensions, amount or density of the different tree components is generally directly correlated with an increase in the total aboveground biomass. That is, as the total biomass increases in value, lesser canopy penetration is expected from the forward radiation. With L-band, the amount of biomass at which the energy no longer passes through the canopy is called the saturation level for that band. Since type of polarization also exerts influence on the penetration depth, the same band with different polarizations will also have different of saturation. These influences are illustrated in Figure 1.
Point be in Figure 1 corresponds to the amount of biomass where the sensitivity of both the L-HH and LHH + C-HV backscattering to biomass measurements becomes saturated. The saturation point for C-HV is represented by point a. The flattening of the lines beyond these points indicates that the backscattering coefficients no longer respond to an increase in biomass of the forest components. Thus, any derived relationship between biomass and backscatter would no longer be valid.
This notion of saturation has important implications for the application of radar to forest areas of high biomass. Returning to figure 1, it can be deduced that equation (5) is no longer applicatibe to biomass levels to the right to b. A possible solution for the estimation of biomass at ranges beyond the soL-HH + soC-HV saturation point is the inclusion of the backscattering from a band with a longer wavelength. Equation (5) could then be modified to
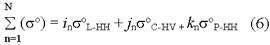
where k is the unit vector corresponding to P-band with HH polarization backscattering coefficient, soP-HV.
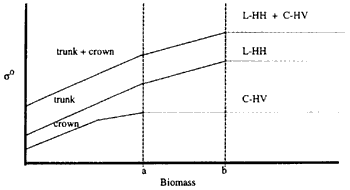
Figure 1. Sensitivities to forest components and theoretical saturation levels for C-HV, L-HH, and LHH + C-HV
2.2 Stand morphology/structure determination
In this study, two general categories are used to differentiate stands according to morphology: those composed of needle-leaved pines/conifers and broad-leaved deciduous/evergreen trees. Pines/conifers are characterized by a pattern where the bole outgrows the lateral branches thus resulting in a well-defined cylindrical trunk and a usually conical crown. The lateral branches thus resulting in a well -defined cylindrical trunk and a usually conical crown. The lateral branches extend up to the relatively lower portion of the central stem producing a narrow and deep crown but the bole is visible almost throughout the entire tree height due to the small size and density of the branches and the needle-lke leaves. Broad-leaved trees are characterized by a less-pronounced trunk but bigger and more voluminous leaves, as well as a wider and thicker crown compared that of the conifers. The lateral branches grow at the same rate, or even faster, than the bole and the definition of the bole can be completely lost due to repeated forking (Dobson et al., 1995a).
As radar backscattering behaviour is directly influenced by he geometric properties of the target, it follows that backscattering from stands of varying structural attributes will be relatively different. A stand of needle-leaved trees, with the small size and density f the leaves and branches as well as the long trunks of the trees, will have a strong L-HH return. On the other hand, the greater attenuation imposed by the bigger and more dense foliage components, coupled with the less conspicuous trunks, will results in a lower L-HH but higher C-HV backscatter from stands of broad-leaved trees. This change could also be related to, say, bit-trunked trees and mallee or scrub type vegetation, or mangroves and trees with more conspicuous trunks. This theory is illustrated in Figure 2.
It is suggested that the above theory can be useful in the determination of tree structure. Given that the sensitivity of the L-HH and C-HV so is a is a function of the trunk and crown components, respectively, then the true ratio of these two backscattering coefficients is a possible measure of tree morphology. This could be illustrated by expression
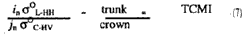
Where TCMI is the Trunk-Canopy Morphology Index. A high TCMI implies a tree structure with more trunk and less crown component while a low TCMI indicates the opposite.
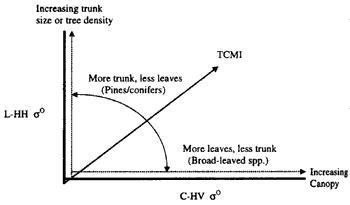
Figure 2. Sensitivity of L-HH and C-HH s° to stand structure properties
3. Application of Radar Theory to Published Research
The effectiveness of the earlier presented theories, models and indices in providing measures of forest aboveground biomass and stand structure is assessed and demonstrated by applying them to actual and modeled data published related works of other investigators. As most of the data and results from the published papers are reported in the form of graphs, interpolation was done to obtain measurements for the backscattering coefficients and the corresponding forest biomass levels.
3.1 Data Sources
Listed and individually described in Table 1 are the different studies used in the assessment of the theories and concepts previously presented. All but two of the studies involved the use of either SIR -C/S-AR or AIRSAR system. In all of these, the HV and VH polarizations were regarded as identical and only the HV data were hence included in the analysis. The incidence angles utilized ranged from 19°-50° and varying but closely related saturation limits were observed.
3.2 Radar backscatter versus forest biomass
All of the studies considered here, and most of the research so afar reported in the literature, have correlated radar backscatter with forest biomass using single radar wavelength and polarization combinations. Although some of the forests investigated are composed of broad-leaved and mixed broad-leaved/needle-leaved species, the majority of the sites are vegetated by managed, even-aged and mono-specific coniferous stands located in temperate zones. Due to known difficulties associated with rugged topography, the different forests areas selected as study sites are located in flat to moderately-sloping areas.
In the interpolation of data from the different studies, only the L-HV backscattering dta were taken into account owing to the concepts discussed earlier. This qualification limited the number of cases considered, as only a few of the related investigations have used either or both of the L-HV and C-HV backscattering data. A maximum of 300 tons/ha was used in view of the typical saturation of radar measurements at all bands beyond this biomass level. For both L-HH and C-HV, so was observed to increase linearly with biomas until the saturation level for each of these wavelength-polarization combinations is reached. The TCB and TCM indices were computed based on the intensity (or true) values of the individual backscattering data i.e. not decibels. The so values were converted into intensity values through the formula
X = - log (s° /10) (8)
where x is intensity or true value and s° the backscattering coefficient (in decibel). No significant variation was observed between the TCBI and TCMI values within the same stand structure category from the different investigations. The similarity in the values of these indices occurred over the entire biomass range, except for some cases at low biomass levels, where inconsistent results were obtained. Factors such as strong influence of terrain/ground conditions on the radar backscatter due to low stand/crown component volume and density, may have caused these results. In general, at biomas levels below the radar saturation limits, positive linear relationships between total aboveground biomass and the two indices were observed. In comparing these index values, the effects on the data of factors such as dissimilarities in the radar systems used (e.g. viewing angle and platform altitude), discrepancies in the applies calibration methods, terrain variations, stand interpolation of the backscattering data and biomass levels, should however be noted.
The correlation between forest biomass and L-HH, and TCBI are presented in Table 2.
3.2.1 TCBI and total forest aboveground biomass
Table 2 indicates that in the case of C-HV, the use of TCBI (L-HH + C-HV) brought about a better correlation between radar backscatter and total forest biomass. On the other hand, in the case of L-HH, mixed results were obtained. Although much is still to be desired regarding both the number of cases and data considered, this finding is generally in support of the theory presented above (see equation (5)). Following are biomass equations derived based on linear regression analysis performed on the TCBI and biomass data from the various studies.
| Autors/s | Radar System | Band/s used | Polarization | Incidence Angle |
Study Sites/s | Dominant Vegetation | Saturation Limits(ton/ha) | Remarks | |
| 1. | Imhoff, 1993 | JPL SAR(AIRSAR) | P-,L-,C- | HH,VV,HV | 40° -50° | a)Hawaii b)North America & Europe | a)Tropical broad-leaved
evergreen b)Conifers |
C-=20 L-=40 P-=100 |
HV and VHTreated as the same |
| 2. | Souyris et al., 1995 | SIR-C/X-SAR | a)L-,C- b)X- |
a)HH,VV,HV b)HH,VV |
26.4° | Less Landes Forest, France |
Pines/conifers | 50 | -do- |
| 3. | Dobson et al., 1995a | SIR-C/X-SAR | a)L-,C- b)X- |
a)HH,VV,HV b)HH,VV |
31° | Raco Supersite, Michigan, USA | Broad-leavedHardwoods and conifers | (multi-step approach accurate up to 250 t/ha) | -do- |
| 4. | Dobson et al., 1995b | SIR-C/X-SAR | L- | HH,VV,HV | 19° -47° | Raco Supersite, Michigan, USA | Broad-leaved hardwoods & conifers | - | -do- |
| 5. | Hsu et al., 1993 | AIRSAR | P-,L-,C- | HH,VV,HV | 45° | Les landes Forest, France | Pines/conifers | - | -do- |
| 6. | Dobson et al., 1992 | AIRSAR | P-,L-,C- | HH,VV,HV | 40° -50° | a)Les Landes forest, France b)Duke forest, USA |
Pines/conifers | P- = 200 L-=100 |
-do- |
| 7. | Karam et al., 1995 | AIRSAR | P-,L-,C- | HH,VV,HV | Pines/conifers | - | -do- | ||
| 8. | Harrel et al., 1995 | JERS-1 &ERS-1 | L- & C- | HH & VV | 45° | Les Landes forest, France | Pines/conifers(White and black spruce) | - | |
| 9. | Christensen et al. 1990 | AirborneSAR (P-3) | L- | HH,VV, | 42° -49° | Duke Forest, USA | Pines/conifers | 100 | |
| 10. | Moghaddam et al., 1994 | AIRSAR | P-,L-,C- | HH,VV,HV | 35° -50° | Maine, USA | Mixed hardwoods and conifers | 200 (for 40°-50°) 300 (for °35) 150 (for 50°) |
L-HH data Not available from report |
| 11. | Ranson & Sun, 1994 | AIRSAR | P-,L-,C- | HH,VV,HV | 25°,35°,50° | Maine, USA | Mixed hardwoods and conifers | 157 | L-HH datanot available |
| 12. | Ranson et al., 1995 | AIRSAR | P-,L-,C- | HH,VV,HV | 25°,35°,50° | Maine, USA | Mixed hardwoods and conifers | C-=100, L-=200P-=>200 | C-HV data not available |
| INVESTIGATORS | SAR Backscatter | CORRELATION COEFICIENT | |||
| PINE/CONIFER | BROADLEAF | ||||
| Biomass (tons/ha) | Biomass (tons/ha) | ||||
| 0-300 | 20 - 150 | 0 - 300 | 20 - 150 | ||
| 1.IMHOFF,M.L.(1993) | L-HH | 0.7354 | 0.3140 | 0.8219 | 0.9321 |
| C-HV | 0.6378 | 0.0051 | 0.6801 | 0.7926 | |
| L-HH+C-HV | 0.6579 | 0.3002 | 0.8134 | 0.9326 | |
| 2.KARAM,et al.(1995) | L-HH | 0.7004 | 0.6347 | ||
| C-HV | -0.0369 | -0.2083 | |||
| L-HH+C-HV | 0.5990 | 0.5571 | |||
| 3.HSU,C.C.et al. (1993) | L-HH | 0.8086 | 0.7581 | ||
| C-HV | 0.1037 | 0.2417 | |||
| L-HH+C-HV | 0.6792 | 0.6131 | |||
| 4.SOUYRIS,et al. (1995) | L-HH | 0.7907 | 0.8764 | ||
| C-HV | 0.4919 | -0.0020 | |||
| L-HH+C-HV | 0.8264 | 0.8289 | |||
| 5.DOBSON,et al.(1992) | L-HH | 0.8731 | 0.9705 | ||
| C-HV | 0.7952 | 0.4900 | |||
| L-HH+C-HV | 0.8769 | 0.8996 | |||
| 6.DOBSON,et al.(1995a) | L-HH | 0.4037 | 0.8322 | 0.0182 | 0.2317 |
| C-HV | 0.7952 | 0.4900 | 0.4431 | ||
| L-HH+C-HV | 0.8769 | 0.8996 | 0.5345 | 0.7759 | |
| 7.DOBSON,et al.(1995b) | L-HH | 0.7999 | |||
| 8.HARISTENSEN dt al. (1990) | L-HH | 0.9888 | 0.9879 | ||
| 9.CHRISTENSEN et al.(1990) | L-HH | 0.9665 | 0.9659 | ||
| 10.MOGHADDAM,et al.('94) | C-HV* | 0.9744 | |||
| 11.RANSON & SUN (1994) | C-HV* | 0.5113 | 0.6393 | ||
| 12. RANSON et al. (1995) | L-HH* | 0.8544 | 0.9557 | ||
Table 2. Correlation between forest biomass and L-HH, C-HV, and L-HH + C-HV backscatter
Pines/Conifers
Imhoff, 1993 : Biomass = 866.75 TCBI - 89.69 (9)
r2= 0.4329
Karam et al., 1995: Biomass = 873.28 TCBI - 78.78 (10)
r2= 0.3588
Hsu et al., 1993 : Biomass = 1476.90 TCBI - 131.00 (11)
Souyris et al., 1995 : Biomass =2609.60 TCBI - 323.09 (12)
r2= 0.6835
Dobson et al., 192 : Biomass = 1495.00 TCBI - 209.59 (13)
r2 = 0.7689
Dobson et al., 1995a : Biomass = 1176.20 TCBI - 22.57 (14)
r2 = 0.1866
BROAD-LEAVED STANDS
Imhoff, 1993 : Biomass = 973.50 TCBI - 74.48 (15)
r2=0.6616
Dobson et al., 195a: Biomass = 5396.50 TCBI - 455.58 (16)
r2= 0.2857
While the regression equation for pines/conifers generated from the results of results of Souyris et al. (1995) had a somewhat higher multiplicative coefficient, similarities could be observed between the equations from Imhoff (1993) and Karam et al. (1995), and from Hsu et al. (1993), Dobson et al. (1992) and Dobson et al. (1995a). The discrepancies between these groups of equations could be attributed to radar system differences and other reasons specified in Section 3.2. The differences in the equations generated for broad-leaved stands from Imhoff (1993) and Dobson et al. (1995a) are significant. However, it should be noted that the latter equation was based on four biomass values only. The highest correlation with biomass was exhibited by that from Dobson et al. (1992), with the equation explaining 77% of the biomass variation.
Given in Table 3 are the TCBI values averaged over the entire biomass range for each of the studies. A comparision of the average biomass between coniferous stands from the different studies, and between coniferous and broad-leaved stands, could be made by using these values as inputs to the corresponding biomass formulas given in equations (9) to (16).
It can be discerned from the table below that the TCBI values from the results of Imbhoff (1993) and Dobson et al. (1995a) for pines/conifers and broad-leaved stands are nearly identical. This was most probably caused by the averaging process, which took into consideration the extreme values within the 0-300 tons/ha biomass range. Moreover, in the case of Dobson et al. (1995a), the insufficiency of available data may also have contributed to the similarity of the averaged results.
| Authors/s | Average TCBI Value | |
| Pines/conifers | Broad-leaved stands | |
| 1. Imhoff, 1993 | 0.2224 | 0.2187 |
| 2. Karam et al., 1995 | 0.1761 | |
| 3. Hsu et al., 1993 | 0.1426 | |
| 4. Souyris et al., 1995 | 0.1514 | |
| 5. Dobson et al., 1992 | 0.2242 | |
| 6. Dobson et al., 1995a | 0.1106 | 0.1125 |
3.3 Radar backscatter versus forest stand structure
Radar backscatter is mainly influenced by the geometric properties of the target. As stated above, forest stands with the same biomass but dissimilar morphology may produce different backscatter readings. Hence, radar data-based equations to estimate total aboveground biomass should be tailored according to the general structure of the forest stand. Although the importance of determining the stand structure prior to generating radar-derived estimates of forest biomass was emphasized in some of the studies (e.g. Imhoff, 1993; Dobson et al., 1995a), not one of the investigations reviewed here have considered stand structure determination through the use of radar backscattering data. In the absence of a priori information on stand structure, radar data-based techniques, such as the application of TCMI, is deemed essential.
3.3.1. TCMI and stand structure
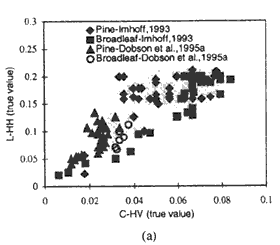
The application of TCMI, which is the true ratio between the L-HH and C-HV backscatter, as a possible measure of stand morphology is premised on the differences in the sensitivity of the two wavelength-polarization combinations to the various tree components. Higher TCMI values are expected for conifers given their bigger trunk and smaller crown component volume compared to their broad-leaved counterpart. This theory has been proven to be generally true based on the test applied using interpolated backscatter and biomass values from related investigations albeit an overlapping of some of the values was observed. The overlaps between the conifer and broad-leaved stand TCMI points, as can be seen from Figure 3, occur at the lower and higher part of the biomass range. This is quite expected due to the minor differences in vegetation structure at low biomass levels and the saturation of radar measurements at high amounts of biomass. The usefulness of TCMI is thus at its optimum when there is a distinct difference in structure between the broad-leaved and needle-leaved trees and the backscattering data are taken at biomass levels below the radar saturation limits.
To further illustrate the usefulness of the TCMI in accounting for the difference in structure between broad-leaved and needle-leaved stand, given in Figure 3b are the graphs corresponding to those in figure 3a but with the L-HH and C-HV data limited to those taken within a biomass range of 20 to 150 tons/ha. By adjusting the lower and higher ends of the biomass range to these levels, the probability that broad-leaved and needle-leaved trees are of the biomass range to these levels, the probability the broad-leaved and needle-leaved trees are more structurally defined, and that the radar measurements are less affected by structurally limits, is increased - and so is the effectiveness of the TCMI. As can be discerned from the figure, the overlapping TCMI points which exist in Figure 3a have been eliminated in the case of Dobson (1995a) and were greatly reduced in the case of Imhoff (1993). For a similar purpose, given below are the average TCMI values within the 0 to 300 tons/ha and 20 to 150 tons/ha biomass range for the different studies. The 20 to 150 tons/ha range caused a higher dynamic range between the TCMI values, and hence a better separation, of the pine and broad-leaved stands. Interestingly, the average TCMI values generated from the different investigations for pines/conifers, except those form Hsu et al. (1993), are similar though the data were taken from different study sites and conditions. The same observation holds true in the case of the values from the broad-leaved stands.
| Autors/s | Average TCMI Value | |||
| Pines/conifers | Broad-leaved stands | |||
| 0-300 | 20-150 | 0.300 | 20-150 | |
| 1. Imhoff, 1993 | 3.3695 | 3.7013 | 2.5168 | 2.3430 |
| 2. karam et al., 1995 | 3.2902 | 3.4119 | ||
| 3. hsu et al., 1993 | 5.4144 | 5.5431 | ||
| 4. Souyris et al., 1995 | 3.6145 | 3.2267 | ||
| 5.Dobson et al., 1992 | 3.4657 | 3.5937 | ||
| 6. Dobson et al., 1995a | 3.7642 | 3.9058 | 2.5882 | 2.5119 |
4. Summary and Conclusions
The capability of microwave energy to penetrate forest vegetation makes possible the extraction of information on both the foliar and woody components from radar data. Depth of penetration, and hence the type of derivable information, is dependent upon parameters relating to both the radar sensor and the target such as the wavelength, polarization and incidence angle used, as well as the geometric and dielectric properties of the target vegetation.
 |
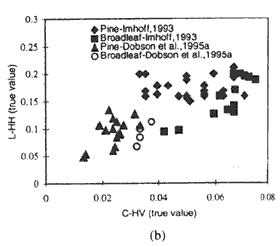 |
| Fig. 3 TCMI values within 0-300 tons/ha (a) and 20 - 150 tons/ha (b) biomass ranges based on interpolated data from Imhoff, 1993 and Dobson et al. 1995a. | |
At C-band, the backscattered energy is correlated mainly with the crown constituents such as the leaves, twigs and small branches. Information on the other components beneath the canopy can be sensed through the use of bands with longer wavelengths such as the L-or P-band. The sensitivity of co-polarized and cross-polarized waves to the shapes and orientation of the different tree constituents provide an added advantage in the information extraction procedure. Given the relatively greater degree of penetration by horizontally-polarized waves and the strong interaction of the vertically-polarized energy with the vertically-oriented canopy parts, different wavelength-polarization combinations can be hen used to suit the purpose of the study. In the field of forest biomass estimation, the Trunk-Canopy Biomass Index (TCBI), which is the sum of the L-HH and C-HV backscatter, can be a measure of the total aboveground biomass as both the crown and trunk layers are taken into consideration. The relationship between TCBI and biomass, however is not expected to be unique for a whole forest vegetation owing to possible variations in morphological structure of to be unique for a whole forest vegetation owing to possible variation in morphlogica structure of the stands within the area. It is important therefore that stand structure be first considered to allow a more accurate biomass assessment by the TCBI parameter.
In the this study, two categories are used to classify stands according to structure: the needle-leaved pines/conifers and the broad-leaved deciduous/evergreen trees. Under the first categorgy are trees having a long main stem and a usually deep, narrow and cone-shaped crown composed of needles and small branches. Trees under the latter have a less-pronounced trunk, a wider and thicker crown with more voluminous leaves. Consequently, an index of the relative proportions of the crown and the trunk may be indicative of the approximate tree morphology. It is believed that the Trunk-Canopy Morphology Index (TCMI), which is the raito of the L-HH to C-HV backscatter, provides a measure of tree structure.
A two-stage procedure for forest biomass estimation is therefore proposed. The first stage involves the determination of the stand structure category based on the TCMI. Once the structure is known, a specific structure-dependent TCBI could then be applied for the biomass estimation process.
The assessment of the effectiveness of the introduced theories carried out based on interpolated date from published research produced promising results. TCMI has been found to provide a good measure of the general stand structure which is deemed necessary prior to the application of biomass estimation techniques. The overlapping of the TCMP values of the needle-leaved and broad-leaved stands which occurred at both ends of the forest biomass range could be explained by the near homogeneity in structure of the stands at low biomass levels and the saturation of radar measurements at high amounts of biomass. In terms of biomass estimation, a higher correlation between TCBI and forest biomass was achieved compared to that of C-HV in all of the studies considered but mixed results were obtained in the case of L-HH. Comparison of the results is made difficult by the limitations both in the number of related studies reported in the literature and in the amount of data available from each of the studies, inherent difference in the radar systems used, differences in he applied calibration techniques, in-situ variations (i.e., in terms of stand structure and composition, topography and other terrain features) and possible interpolation-related errors. Nevertheless, given all of these limitations, the results support the theories proposed.
To further verify the validity of the results, subsequent application of the TCB and TCM indices using extensive field data and AIRSAR images of an independent study site in the edge of the Blue Mountains National Park in New South Wales, Australia will be made.
References
- Dobson, M.C., F.T. Ulaby, L.E. Pierce, T.L. Sharik, K.M. Bergen, J. Kellndorfer, J.R. Kendra, E.Li, Y.C. Lin, A. Nashashibi, K. Sarabandi and P. Siqueira, 1995a. Estimation of forest biophysical characteristics in Northern Michigan with SIR-C/X-SAR.IEEE Transactions on Geoscience and Remote Sensing 33(4): 877-894.
- Dobson, M.C., F.T. Ulaby, T.Le Toan, A. Beaudoin, E.S. Kasischke, and N. Christensen, 1992. Dependence of radar backscatter on coniferous forest biomas. IEEE Transactions on Geoscience and Remote Sensing 30(2): 412-415.
- European Space Agency, 1995. Satellite radar in agriculture: experience with ERS-1, ESA SP-1185, ESA Publications Division, Noordwijk, The Netherlands, 71 p.
- Hsu, C.C., R.T. Shin, J.A. Kong, A. Beaudoin, and T.Le Toan, 1993. Application of theoretical model for microwave remote sensing of forest. Proceedings of the International Geoscience and Remote Sensing symposium, Tokyo, Japan, 18-21 August, 2:295-597.
- Imhoff, M.L., 1993 Radar backscatter/biomass saturation: observations and implications for global biomass assessment. Proceedings of the International Geoscience and Remote Sensing Symposium. Tokyo, Japan, 18-21 August, 1:43-45.
- Karam, M.A. F. Amar, A.K. Fung, E. Mougin, A. Lopes, D.M. Le Vine, and A. Beaudoin, 1995. A microwave polarimetric scattering model for forest canopies based on vector radiative transfer theory, Remote Sensing of Environment 53:16-30.
- Souyris, J.C., and T.Le Toan, 1995.Inversion of Landes forest biomass using SIR-C/X-SAR data: experiment and theory. Proceedings of the International Geoscience and Remote Sensing Symposium, Florence, Italy, 2:1201-1203.
- UNSW Centre for Remote Sensing, 1996. Microwave Remote Sensing Course Notes, First Sections, School of Geomatic Engineering, The Univ. of New South Wales, Kensington, Australia.